|
Time has flown by at OIMB! I can't believe our last week is coming to a close. It's been an amazing experience and I can't believe it's almost over. This week in lab has been a whirlwind; I've been trying to test out my 'Cat' and 'Dog' probes with varied degrees of success. My 'Dog' construct seems to be fairly unstable and expresses inconsistently in both starfish and barnacles with little localization. My 'Cat' construct, on the other hand, appeared to cause high levels of Rho activation, especially during second meiosis. However, since I injected the 'Cat' probe along with a cocktail of other probes, I couldn't see the 'Cat' expression alone. Instead, I had to judge whether other probes had a reaction to my probe (almost as if my probe was a treatment, not a fluorescent tag). Ultimately, more experiments are needed to better understand whether the probe is activating myosin and promoting contractility. Although I won't be here to help, hopefully my probes produce useful results in future experiments.  Hiking some of the South Slough trails with Renee and Megan! We are having our final poster session this week as well. Although it was definitely a challenge to integrate everything I worked on this summer into one cohesive final product, I'm fairly happy with how it turned out. Despite my ‘Cat’ and ‘Dog’ probes not quite working as planned, my poster is a reminder that I did have some success with other probes this summer and did get some interesting results.  I'm going to miss juggling oocytes between microscopes, babysitting eggs during long recording sessions on the confocal, poking at nudibranchs in the sea tables, bonfires at the OIMB beach, and most of all, the people I've met. We've formed a tight-knit community here at OIMB, and it's definitely going to be hard to leave. Thank you to everyone who helped make this summer so special!
0 Comments
After testing out some of my probes last week and determining that they are lethal at high concentrations, I took a different approach to injecting them in hopes of more informative data. First, I injected the probes at lower concentrations. This experiment produced more informative results; I found that although the oocytes still don’t go through anything resembling meiosis, they don’t explode (as often). Additionally, as expected, oocytes experienced high Rho activation throughout, almost as though I had unplugged something important that was inactivating Rho before… This period of high Rho activation made the cells turn completely white, since the Rho probe lit up all over the cortex. This led to the mutant's current nickname: 'whiteout'. As mentioned above, unfortunately, not all oocytes remained unharmed during the duration of my experiments. After addition of hormone to those oocytes injected with high concentrations of either my PBR or Snowflake-PBR probes, I found that the oocytes would try to contract as if they were trying to start meiosis, but would fail quite dramatically. A video is attached – warning, it’s a little gory! (sorry for the low quality)
Apart from accidentally exploding some oocytes this week, I have made good progress on other aspects of my project. My ‘dog’ (Rock regulatory domain) is almost ready for injection, with the ‘cat’ (Rock kinase domain) not far behind. Also, the pieces (front, middle, and tail) of the S. purpuratus Rock are ready for Gibson assembly: a cloning method that allows for assembly of multiple small fragments into a single cohesive sequence. This will allow us to finally construct a probe specific to an echinoderm Rock. Although I probably will not have a chance to include ‘cat’, ‘dog’, and the S. purpuratus data on my poster I will still try out these probes before I leave in hopes of confirming my preliminary results.
Outside of lab, OIMB had its notorious ‘Spineless Soiree’ (otherwise known as the invertebrate ball) today. We all dressed up as our favorite marine invertebrates and strutted down a catwalk in the middle of our dining hall. I went as the purple urchin, S. purpuratus, along with Renee and Megan. We all carefully crafted urchin spines out of purple wrapping paper and huddled together with our spines in all directions. We also brought a little lantern to represent Aristotle's lantern. Although it was hot, messy, and pretty ridiculous, it was still a lot of fun!
I’ve had quite an exciting week in lab; the time finally came to test out some of my probes! I tried out the two Ect2 mutants I was making last week that have mutations in the regions that are thought to bind Rock (affectionately termed Putative Binding Regions, or PBR). In one version of the mutant only the Rock binding regions are mutated (simply PBR), but I also created a version of the mutant that contains both the Rock mutation and the Snowflake mutation I documented in week 3 of this blog (Snowflake-PBR). I started injecting oocytes with both versions of the probe alongside wild-type Ect2 and Snowflake Ect2 controls.
Left: a bat star with a protruding stomach (a sign of a healthy animal and ripe ovaries!) Right: a starfish without a protruding stomach. Unfortunately, the oocytes I have injected so far have been largely uncooperative. They did not express either probe at a high enough level to image, and many of them seemed quite sickly. The ones injected with the PBR probe also had odd zones of bright green Rho activity and behaved very strangely when I added hormone. Despite this oddity, they had dim Ect2 expression, which was a little discouraging. The snowflake-PBR mutants also had somewhat dim expression, but I have hope that the batch of oocytes is at fault, not the batch of Ect2 mutants.  One of our dissecting microscopes that we use to sort embryos and look at eggs, larvae, jellyfish, and lots of other cool things! We will be taking this microscope with us to the Charleston Marine Life Center to educate the public about our research. Aside from injecting and imaging a number of oocytes this week, I have been getting ready for presenting my project at the Charleston Marine Life Center this weekend. I fertilized some of my bat star eggs, so hopefully I will have some larvae by Saturday that little kids can look at under a dissecting microscope. I am looking forward to talking to the general public about my research, but a little concerned that my research is difficult to explain to kids. Hopefully they will appreciate looking at movies and larvae as much as I do!
Time has been flying at OIMB! This week has been particularly busy for me, although not with injections and imaging this time. I’ve been working on the molecular biology aspects of my project, specifically in terms of mutating Rock binding sites in Ect2, amplifying full length bat star Rock, and attempting to stitch together a purple urchin Rock.
The purple urchin Rock has been as prickly and unyielding as the urchins themselves; after many failed PCR attempts, we decided to abandon hope of obtaining the full sequence in one go. Instead, we will combine the pieces that we have already obtained using a technique called Gibson assembly. Additionally, we will tag the N-terminal (front half) kinase domain with mCherry (a red fluorescent protein) and the C-terminal (tail half) regulatory domain with GFP. Both of these constructs may be valuable despite being separated from their other halves; naturally, the regulatory domain is constantly chasing the active kinase domain, so we have been referring to the kinase domain as the ‘cat’, and the regulatory domain as the ‘dog’.
A few of this week's sunsets. Left: sunset at Bastendorff Beach, right: sunset at the OIMB beach. In addition to battling the purple urchin Rock, we have been working hard on mutating the Rock binding sites in Ect2. After using our gene blocks to mutate the Rock binding sites and amplify the new version of Ect2, we transformed the purified PCR product into E. coli and let the little bacteria make many copies of the plasmids we want. We then extracted the plasmids and conducted several digests with restriction enzymes that cut in known locations to confirm that the E. coli actually amplified what we wanted. Although the digests looked a little weird, we sent some of the purified plasmid DNA for sequencing to verify the presence of our constructs. Despite the setbacks I've described, life inside and outside of the lab has been fun. The REUs took a weekend trip up to Newport last weekend to visit the Oregon Coast Aquarium (see photos above). Watching the rays and skates swim above me in the "Passages of the Deep" tunnels was magical, as was watching the octopus feeding at the Hatfield Marine Science Center. I look forward to continuing exploring the Oregon coast and hopefully getting back some sequences this weekend to start injecting my probes next week!
This has been quite a busy (and rewarding!) week in the lab. The gBlocks (artificially created double stranded DNA fragments) I was waiting for to construct the Ect2 mutant finally came in the mail. I started creating the mutant by using the gBlocks (which have the mutations in the Ect2 binding sites present in the sequence) as primers for PCR amplification of wild type Ect2. This allows me to introduce the mutations I want in the Ect2 sequence.
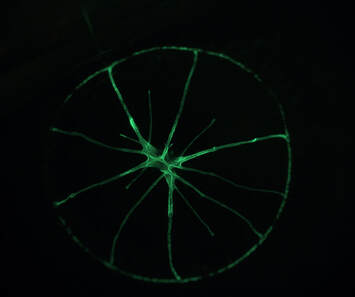
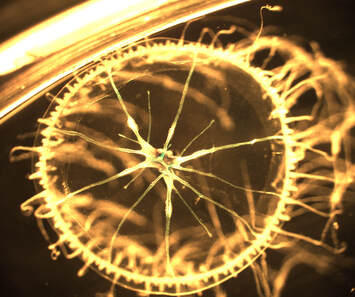
Naturally fluorescent jellyfish under a dissecting microscope. These jellyfish are actually quite small and some individuals naturally fluoresce under a green laser. The left is just the green laser, the right is both normal transmitted light as well as the laser (which is why you can see some green residue towards the center). In preparation for looking at my Ect2 mutant, I injected some sea star oocytes with other mutated versions of Ect2 to provide a basis for comparison. In addition to T808A or Snowflake, a mutant I have previously posted a video of, I also injected a mutant named Silent Spring (because of the presence of ‘DDT’ in its amino acid sequence), as well as a similar mutant named Snow Leopard. These mutants both interfere with binding sites for a protein called Cdk1, a cell cycle regulator which phosphorylates Ect2.
On the Rock front, we received the sequences from the attempted S. purpuratus Rock amplification. For the most part, the sequences looked really clean. We successfully cloned both the front and back ends of the Rock gene into the appropriate vector. The next step is to amplify the missing middle section and combine the middle section with the front and tail to create a complete version of the gene. Additionally, we now have the Patiria miniata putative Rock sequence. After designing primers for the P. miniata Rock, Kara managed to PCR amplify the gene on the first try! This means that I could potentially make another version of the probe using the putative P. miniata Rock, which is exciting!
This week, I continued my pursuit of the elusive Rock gene. I am trying two approaches to tag and follow Rock: the first is to use a Rock probe originally designed for Xenopus laevis (African clawed frog), and the other is to design an echinoderm-specific Rock probe.  An X-ray of my model organism, Patiria miniata. Photo credit to Edward Nichols and the Hanson-Meekins Animal Hospital. While struggling with the PCR of the echinoderm Rock sequence, I tried injecting sea star oocytes with various concentrations of frog Rock. Unfortunately, this also presented several obstacles; since Rock is a fairly large protein (about 4 kilo base pairs) and maybe easily degraded, it is very tricky to actually see its activity within the cell. However, I did get one successful movie of Rock making waves on the cell surface that appear to overlap and immediately follow Rho waves. Although the Rock fluorescence is fairly dim, the movie is long enough to create a kymograph to further analyze wave activity. Additionally, I am continuing to try to alter probe concentrations and length of time after injection to try to catch more Rock activity.  A female (red rock?) crab with extruding eggs bravely caught by Richard. After many unsuccessful runs, we finally obtained PCR product for the full-length Rock gene, as well as the front and back halves separately (thanks to Kara and her perseverance!). After purifying the PCR product, we went ahead with the next steps in making the GFP probe. First, we ligated the PCR product with a GFP vector, thus introducing the PCR product onto a circular plasmid. We then transformed the plasmids into E. coli cells and conducted colony PCR to confirm the transformation success.
Outside of the lab, the REU interns all went camping this weekend at Sunset Bay. After being woken up by sea lions barking, we all went tide pooling in the morning. We were rewarded for waking up for the low tide; we found some giant crabs, sea lemons, huge sea urchin colonies, and even spotted the arms of an octopus as it drifted by.
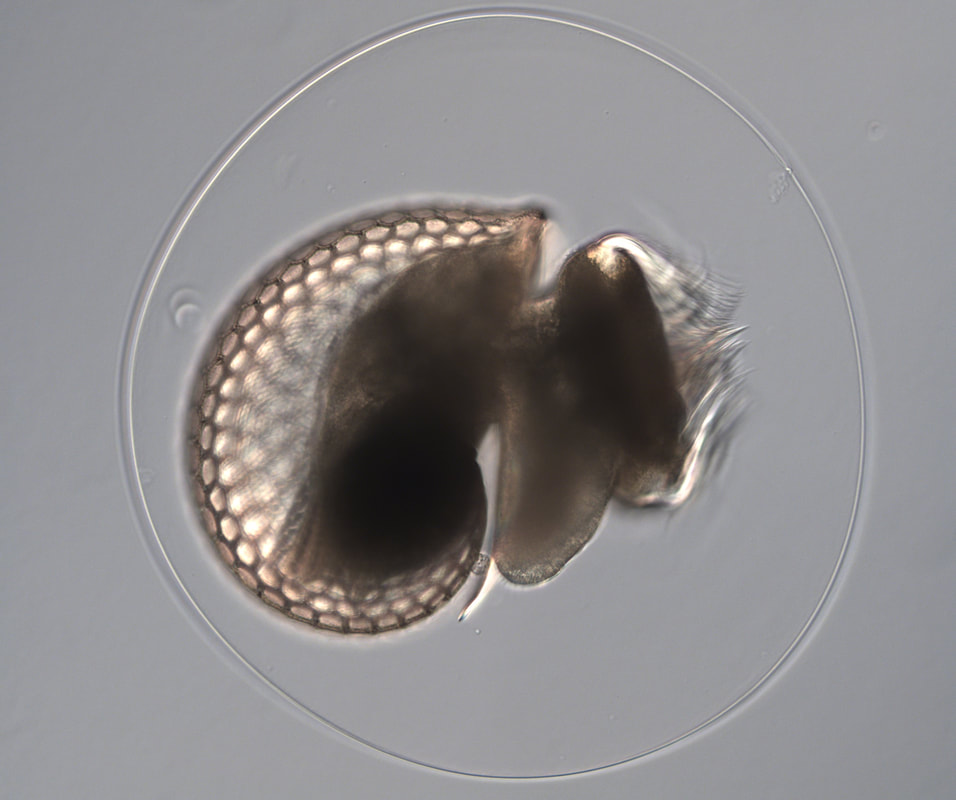
After solidifying my project plan last week, I started working on executing that plan this week, but not without a few unforeseen obstacles. The first step in designing the Rock probe which I hope to inject into sea star oocytes is to use PCR (polymerase chain reaction) to amplify the predicted Rock sequence from the sea urchin genome. Unfortunately, this step has been unsuccessful so far; after trying a handful of different primers (short DNA sequences that initiate replication). However, not all is lost – I am continuing to adjust variables and troubleshoot the process in hopes that the Rock sequence will appear.  The second unforeseen obstacle this week was the uncooperative nature of some of the sea star oocytes I injected. On Monday, I injected fresh sea star oocytes with several different probes (mostly for practice, but also to get a hands-on sense of the wave phenomenon caused by different probes). Although one of the probes did not produce the expected waves, the other, an Ect2 mutant called ‘snowflake’, did cooperate (see kymograph and movie). It exhibited constant expression prior to first meiosis, waves during first meiosis, another period of constant expression prior to second meiosis, and more waves during second meiosis. These waves could be seen by following the fluorescently tagged Ect2. Left: Kymograph of ‘snowflake’ Ect2 wave activity. This kymograph was generated from time lapse movies of wave fronts traveling across the surface of the cell; the top of this image represents the surface preceding first meiosis, and as you look further down the image you can follow the wave fronts during first and second meiosis.
Another surprise I encountered this week came through the mail. Along with the expected ripe sea stars arriving to aid my faulty oocyte problem, the package contained sea slugs! The sea slugs were on their way to a medical center in New York, and are often used for research involving learning and memory due to their large neurons and relatively simple body plan. For now, we have no such lofty goals for the sea slugs, but have been amazed by their ability to produce enormous amounts of eggs – they lay impressive ribbons of thousands of eggs, which have been collecting on the sides of the tank.
A few members of our nudibranch family. Top left: Charlemagne, top right: Chance, bottom left: Chuck, bottom right: no name yet, but open to suggestions!
After orientation to lab and OIMB last week, this week I focused on outlining some of the specifics of my project. I would like to introduce you to my model organism: Pitiria miniata, or the bat star, a type of sea star. Sea stars have beautiful oocytes that are quite large in comparison with other animals and thus are easy to manipulate and interesting to study. The project I have taken on involves targeting a protein known as Rho Kinase (Rock). This protein is hypothesized to be a part of a negative feedback loop involving many other proteins. A variety of proteins need to work in concert in order to regulate cell division, and Rock is one piece of the puzzle. I hope to better understand its activity by designing a fluorescent probe for the sea star Rock protein. This will then allow me to understand and further manipulate the system.
In addition to working on my own project this week, I took a peek at my lab partners amazing projects. Although we all are studying cell signaling during cell division, we all chose different model organisms. Phillip is pioneering the study of cell division in jellyfish eggs and embryos (through a lot of trial and error) and Sadie is working with barnacle embryos. For practice, Sadie injected the embryos with a fluorescent tag that allows for visualization of microtubules, a key component of the cell’s interior that facilitates transport of proteins across the cell.
Click above to download a movie of a barnacle embryo dividing at the 8 cell stage. Apart from all of the incredible projects happening in the lab, I have been exploring the Oregon coast. This weekend, we took a boat ride on the Pluteus along the coast, and although the water was choppy and we weren’t able to collect many animals, the coast was beautiful. Earlier this week, my lab also went tide pooling together, and we collected some of my new favorite animals while traipsing along the beautiful intertidal zone, spotting nests of sea urchins and stumbling across unique invertebrates at every turn. It seems like there is a treat at every turn; while jellyfishing with Phillip on Thursday, we ran into a few harbor seals sunning themselves on the docks. This week has been amazing and I’m looking forward to what else OIMB has in store.
Some adorable harbor seals sunning themselves at the little boat basin.
A few interns finding light at the end of "the tunnel".
Hello! My name is Nina Denne and I just finished my first week as an intern in the REU program at the Oregon Institute of Marine Biology (OIMB). I’m originally from the Chicago suburbs, but I go to Carleton College in Minnesota, where I am majoring in Biology with minors in Russian and Neuroscience. At Carleton, I do cell biology research focused on the bacterium Ralstonia solanacearum, which infects plant tissue and causes bacterial wilt disease. Although I’ve always been fascinated by marine biology, I’ve never had a chance to do any type of marine science before. In fact, I had never even seen the Pacific Ocean before this weekend!
Above: Just a few of the organisms I’ve met this week. Left to right: a sea urchin, a sea star, an anemone, and a jellyfish. At OIMB, I am working in Dr. George von Dassow’s lab. His research focuses on embryology of marine invertebrates including sea urchins, sand dollars, jellyfish, sea stars, and others. Specifically, current research questions involve the role of proteins Ect2 and Rho in the organization of actin (a part of the cell’s cytoskeleton) during cytokinesis (cell division). However, prior to delving into the research, we needed to develop some technical skills. One of the first skills we tackled was injecting developing embryos with probes containing instructions to make green fluorescent protein (GFP) fused to a protein of interest. Additionally, we practiced melting and reshaping glass tubes to form a curved tiny glass tube that will let in a single embryo at a time. This glass tube is connected to rubber tubing with a mouthpiece on the other end; this is essential because transferring embryos requires high precision, and mouth-eye coordination is more precise than hand-eye coordination.
Left: A purple sea urchin releasing its eggs. Right: Injection of recently fertilized sea urchin eggs with a red fluorescent probe. Since most of Dr. von Dassow’s work involves microscopy and working with gametes and embryos, we also tried looking at a variety of subjects under different microscopes for practice. The subjects included sea urchin embryos, sea star oocytes, jellyfish oocytes, and more. One of the subjects I was most excited to work with was the diatom – a single celled alga with a silicon cell wall. Although I had worked with diatom genomes in a previous lab at the Chicago Botanic Garden, I never met them. This week, I was able to see a diverse array of diatoms under the microscope, collected from a plankton tow. In addition to diatoms, I also met some dinoflagellates, a group of protists found in plankton that have a single tail (called a flagellum). The species that Dr. von Dassow keeps as a pet has another interesting quality: they are bioluminescent, meaning they glow in the dark when slightly agitated. On Wednesday evening, I came back to lab in the evening to see the light show produced by the dinoflagellate Pyrocystis and was amazed by waves of glowing green specks!
|

AuthorI am a rising junior at Carleton College, majoring in biology with minors in Neuroscience and Russian. I'm very excited to be working in Dr. George von Dassow's lab this summer, where I will be studying cell biology and embryology of marine invertebrates. Archives
August 2019
Categories |
||||||||||||||||||||||||||||||||||||||



























 RSS Feed
RSS Feed